PLANT VIRUSES
Economic Impact
- Crop plants in most parts of the world are subject to significant losses as a result of infections by viruses and viroids.
- Worldwide losses of approximately 60 billion dollars per year have been reported.
- Crop plants of every type can be stricken by virus diseases.
- 50 million citrus trees have been lost or rendered unproductive as a result of tristeza virus.
- 200 million cocoa trees in parts of western Africa lost to the swollen shoot disease.
- Rice plantations sustain very large losses.
- Barley yellow dwarf virus is one of the most serious pathogens of grain crops.
- Ornamental, vegetable, and field crops are susceptible to potyviruses wherever they grow.
Morphology and Genome
- Plant viruses are morphologically similar to animal viruses, and they have similar types on nucleic acid.
- Many are long, flexible, very narrow helixes, but of a length approaching that of Escherichia coli.
- The Closterovirus (beet yellow) is 1.3 µm in length but very narrow (<10 nm), can not be visualized in a light microscope.
- The capsid of a plant virus is generally composed of a single protein.
- The genome of most plant viruses is composed of either ss or dsRNA.
- Many plant viruses exhibit a replicative cycle similar to the lytic replication cycle of bacteriophages.
Multipartite Plant Viruses
Although multipartite genomes also segmented, each genome segment is packaged into a separate virus particle.Segmented virus genomes are those which are divided into two or more physically separate molecules of nucleic acid, all of which are then packaged into a single virus particle.
Virus having partition strategies are:
| Viral Family | Segment |
| Begomovirus (Geminiviridae) (single-stranded DNA) | Bipartite |
| Comovirus (single-stranded RNA) | Bipartite |
| Furovirus (single-stranded RNA) | Bipartite |
| Tobravirus (single-stranded RNA) | Bipartite |
| Partitiviridae (double-stranded RNA) | Bipartite |
| Bromoviridae (single-stranded RNA) | Tripartite |
| Hordeivirus (single-stranded RNA) | Tripartite |
The Strategy of Genome Segmentation
These discrete particles are structurally similar and may contain the same component proteins, but often differ in size depending on the length of the genome segment packaged. In one sense, multipartite genomes are, of course, segmented, but this is not the strict meaning of these terms as they are used here.
- Genome segmentation reduces the probability of breakages due to shearing, thus increasing the total potential coding capacity of the genome. The disadvantage of this strategy is that all the individual genome segments must be packaged into each virus particle, or the virus will be defective as a result of loss of genetic information.
- Separating the genome segments into different particles (the multipartite strategy) removes the requirement for accurate sorting, but introduces a new problem in that all the discrete virus particles must be taken up by a single host cell to establish a productive infection. This is perhaps the reason multipartite viruses are only found in plants.
- Many of the sources of infection by plant viruses, such as inoculation by sap-sucking insects or after physical damage to tissues, result in a large inoculum of infectious virus particles, providing opportunities for infection of an initial cell by more than one particle.
Taxonomy
At present, plant viruses do not fit easily into a basic taxonomic scheme and are grouped according to the types of diseases they cause.
- There are over 1,000 plant viruses have been reported.
An additional, 200 viruses remain unclassified.
- Geminivirus and Caulimovirus are naked, circular DNA viruses; ss and ds, respectively.
All the other plant viruses are naked,linear ssRNA viruses.
Geminiviridae
Ambisense single-stranded circular DNA genomes and are members of class II of the Baltimore classification of viruses.
The genome can either be a single component of 2500-3000 nucleotides, or two similar-sized components.
They have an elongated, geminate with two incomplete icosahedra joined at the missing vertex.
The capsids range from 18-20 nm in with a length of about 30nm.
Viruses with bipartite genomes (begomoviruses only) have these components separated into two different particles, therefore more than one virus particle is required to infect a cell.
Geminiviruses replicate via a rolling circle mechanism like bacteriophages such as M13, and many plasmids.
Replication occurs within the nucleus of an infected plant cell.
Genera
- Mastrevirus
- Curtovirus
- Begomovirus
- Topocuvirus
These viruses are responsible for a significant amount of crop damage worldwide.
Epidemics of geminivirus diseases have arisen due to a number of factors, including the recombination of different geminiviruses coinfecting a plant, which enables novel, possibly virulent viruses to be developed.
Other contributing factors include the transport of infected plant material to new locations, expansion of agriculture into new growing areas, and the expansion and migration of vectors that can spread the virus from one plant to another.
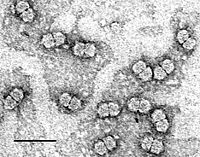
Geminivirus (Size bar indicates 50 nm) Picture by Kassie Kasdorf
Potyviruses
- Potyviruses are the most destructive of plant viruses.
- Over 10% of the plant viruses are potyviruses.
- Most of them have narrow host ranges.
- Most of the world's important vegetable and field crop growing areas sustain serious losses to potyviruses.
- In some cases, mixed infections involving more than one Potyvirus or a Potyvirus and another virus may produce much more serious diseases than are caused by single infections.
Transmission
- No plant virus is known to use a specific cellular receptor of the type that animal and bacterial viruses use to attach to cells.
- After replication in an initial cell, the lack of receptors poses special problems in recruiting new cells to the infection.
- Many plant viruses are transmitted by insects or arthropod vectors and can replicate inside the cells of these vectors.
- Other viruses are transmitted directly from plant to plant.
- Plant virus appear to lack specific mechanisms for penetration of plant hosts.
- They enter as a result of mechanical damage.
Causes of Mechanical Damage
Insect injury
Weather deterioration
Mechanical abuse from cultivation methods
The Role of Vectors in Transmission
- Viruses must enter through wounds or be assisted by other plant parasites, including nematodes, fungi, and, most often, insects that suck sap.
- Viruses infect the mouth parts of insects and are transferred to uninfected host during normal feeding as theinsects suck sap
- Some plant viruses are stored in the foregut of aphids and inoculated into the plant by regurgitation during feeding.
- Some plant hosts, remain symptomless and only serve as reservoirs of infection.
- The wound tumor viruses multiply in the tissue of leafhoppers before they move to the salivary glands.
- Virus is then transmitted by the insect bite.
- The Topovirus (tomato spotted wilt) and the Rhabdovirus (lettuce necrotic yellow) are two other examples of virusesthat infect the insects that transmit them.
Modes of Transmission
Plant Virus |
Mode of Transmission |
| Geminiviruses |
A species of whiteflies, treehoppers |
Cauliflower mosaic virus |
Aphids |
Bean mosaic virus |
Wounds |
Tobamovirus |
Pollen |
Bymovirus |
Fungi |
Potyvirus |
Aphids |
Rymovirus |
Mites |
Potato yellow dwarf virus |
Leafhoppers and aphids |
Wound tumor virus |
Leafhoppers |
Tobacco necrosis |
Parasitic fungi |
Tobacco ringspot virus |
Nematodes |
Note: Specific proteins from virus capsids interact with proteins from vector-virus systems (nematodes, fungal, aphids, mites, etc.)
==> The most important agents of transmission are sucking insects.
The Intracellular Spread of Virus
Movement Proteins
-
Plant viruses encode special movement proteins that spread the virus from cell to cell through the plasmodesmata.
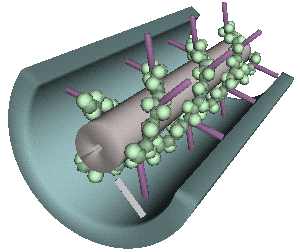
- Plasmodesmata channel surrounded by proteins
Source: MicrobiologyBytes.com
- These channels of communication are too small to allow for the passage of virus particles or genomic nucleic acids.
- Many (if not all) plant viruses have evolved specialized movement proteins which modify the plasmodesmata.
- TMV encodes only four proteins, but one of them is a movement protein (30k).
- CPMV, uses the 58/48k proteins to form tubular structures allowing the passage of intact virus particles to pass from one cell to another.
- Movement proteins were discovered in the last decade.
- Replace virus's cell-killing genes with genes for a protein of commercial value.
- This technique is used for tricosanthin production, an experimental drug for the control of HIV.
Plant Viral Diseases
- Plant cells are generally protected from disease by an impermeable cell wall
- Viral infections can not be cured.
- Most viral diseases of plants are chronic degenerative diseases.
- Plant cell produce proteases to kill viruses but not interferon as in animal cells.
Symptoms of Viral Disease
- Most plant viruses exhibit great host specificity and cause various symptoms in the host.
- Disruption of plant hormones affecting cell division and elongation.
- Mosaic diseases cause mottling of leaves, yellow spots, blotches, and necrotic lesions mostly on flowers and leaves.
- Variegation on leaves and petals (e.g. in tulips).
- A number of viruses cause leaves to curl and/or turn yellow.
- Leaf discoloration.
- Dwarfing of leaves or excessive branching can result from viral infection.
- Abnormal or deformed root growth.
- Wound tumors can occur on roots or stems.
- Slow, stunted plant growth that reduce seed and herbage yields.
Signs
- Necrosis of cells, caused by direct damage due to virus replication.
- Hypolasia, i.e. localized retarded growth frequentlly leading to mosaicism (the appearance of thinner, yellow areas on the leaves)
- Hyperplasia, which is excessive cell division or the growth of abnormally large cells, resulting in the production of swollen or distorte areas of the plant

An infected plant. Source: MicrobiologyBytes.com
Common Viral Plant Diseases
Causative Viral Agent |
Disease |
Caulimovirus |
Cauliflower mosaic |
Almovirus |
Alfalfa mosaic |
Begomovirus |
Bean golden mosaic virus |
Bromovirus |
Brome mosaic |
Carlavirus |
Carnation latent |
Closterovirus |
Beet yellows |
Comovirus |
Cowpea mosaic |
Cucumovirus |
Cucumber mosaic |
Curtovirus |
Beet curly top virus |
Hordeovirus |
Barley stripe mosaic |
Ilarvirus |
Tobacco streak |
Luteovirus |
Barley yellow dwarf |
Mastrevirus |
Maize streak virus |
Nepovirus |
Tobacco ringspot |
Potexvirus |
Potato X |
Potyvirus |
Potato Y |
Rhabdovirus |
Lettuce necrotic yellow |
Tobamovirus |
Tobacco mosaic |
Tobanecrovirus |
Tobacco necrosis |
Tobravirus |
Tobacco ratttle |
Tombusvirus |
Tomato bushy stunt |
Topocuvirus |
Tomato pseudo-curly virus |
Topovirus |
Tomato spotted wilt |
Tymovirus |
Turnip yellow mosaic |
WMV |
Watermelon mosaic |
Control of Plant Diseases
- Breeding of virus-resistant and virus-free plant strains.
- Developing resistant plants is the method of choice because viruses are often spread in seeds or by grafting.
- Control the insects that spread the virus.
Ironically, some Virus-Infected Plants are often Beautiful!
- Potato plants with leaf roll disease have thickened, healthy-looking leaves.
- Grape vines with Pierce's disease turn particularly beautiful shades or orange and red in the fall.
- Tulip plants with tulip break disease have beautifully variegated flowers.
Cultivation of Plant Viruses
In laboratories, plant viruses are cultured in protoplasts (plant cells with the cell walls removed) and in insect cell cultures.
Viruses can be propagated in vivo by inoculating them directly onto plant tissue.
An Emergent Plant Virus
Rarely, there seems to be an example of an emerging plant virus which has acquired extra genes, and as
- The tomato spotted wilt virus (TSWV), a Bunyavirus, has a very wide plant host range.
- Infects over 600 different species from 70 families.
- In recent decades, the virus has been a major agricultural pest all over the world.
- Its rapid spread has been due to dissemination of its insect vector (the thrip Frankinellia occidentalis) and diseased plant material.
- TSWV undergoes propagative transmission and it has been suggested that it may have acquired an extra gene in the M segment via recombination, either from a plant or from another plant virus.
- This new gene encodes a movement protein, conferring the capacity to infect plants and cause extensive damage.